

(一)T细胞受体

T细胞受体(T cell receptor,TCR或Ti)是T淋巴细胞表面识别外来抗原与自身MHc Ⅰ类抗原(或Ⅱ类抗原)复合物的受体,在同种异体移植中TCR也识别单独的非已的MHC抗原。目前已经证实,TCR在细胞表面与CD3密切结合在一起组成TCR/CD3复合物,TCR识别抗原后刺激信号是通过CD3分子传递的。
1.T细胞受体的类型和结构 TCR中的多肽链是异质性的。根据抗原结构和编码基因不同,已发现有α、β、γ和δ四种多肽链。关于TCR多肽链的结构大多是从分析TCR多肽链cDNA或基因组克隆(genomicclones)而来,编码TCR多肽链的基因属于免疫球蛋白基因超家族成员。成熟TCR肽链分子量在40~60kDa之间。根据TCR中异源双体的组成的不同,TCR可分为以TCRαβ和TCRγδ两种类型。
(1)TCRαβ:CD阳性TCRαβT细胞可识别非已MHCⅡ类抗原(同种异体抗原)或自身MHCⅡ类抗原与加工后抗原的复合物CD8阳性TCr αβT细胞则可识非已MHCⅠ类抗原或自身MHC Ⅰ类抗原与加工后抗原的复合物TCRα链分子量40~50kDa的酸性糖蛋白,β链40~50kDa不带电或碱性糖蛋白。α和β链各由一个可变区(V区)和一个恒定区(C区)组成,与Ig的V区和C区大小相似,属于免疫球蛋白超家族成员。TCRα、β链的V区约含102到109个氨基酸,在V区部分由两个半胱氨酸形成链内二硫键,组成约含50~60氨基酸残基的环肽,这与IgV区结构和功能相似,是特异性识别外来抗原的结构域。TCRα、β链的C区约含138到179个氨基酸,每个C区形成由链内二硫键连接的环肽。α、β链在连接肽(connectingpeptide)形成链间二硫键。穿膜区约由20~24氨基酸组成,α链穿膜区含有带正电的1个赖氨酸和1个精氨酸残基,β链穿膜区含有1个带正电的赖氨酸残基,这些带正电的氨基酸与CD3γ、δ和ε链穿膜区带负电的谷氨酸和/或天冬氨酸形成盐桥,稳定TCR/CD3复合物结构,并与CD3传递信息有关。α、β链胞浆部分只有5~12氨基酸长的尾部(图1-2)。
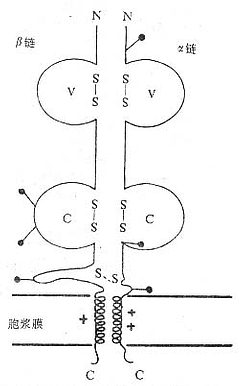
图1-2 TCRαβ异源双体模
(2)TCR γδ:TCRγ和δ链各包括一个Ig样的V区和C区、连接肽、疏水的穿膜区以及一个短的胞浆区尾部,在连接肽区可形成链间的二硫键。γ和δ链的穿膜区各含有1个带正电的赖氨酸,此外δ链还有1个带正电的精氨酸,这些带正电的氨基酸与CD3γ、δ和ε链穿膜区带负电的天冬氨酸或谷氨酸形成盐桥。在氨基酸水平上分析,TCRγ链与β链同源性较高,而TCRδ链与α链同源性较高。在人类TCRγδ有二硫键相连和非共价相连两种形式,而在小鼠只发现二硫键相连的TCRγδ形式。人γ链分子量为36~55kKa,δ链为40~60kDa,γ、δ链的分子量大小取决于多肽骨架的长度和糖基化的程度。
有关TCRα、β、γ、δ链基因的结构和重排见第三章“免疫球蛋白超家族”
2.两种类型TCR T细胞的比较 TCRαβ与TCRγδ不仅组成受体多肽链的结构不同,而且具有这两种类型受体T细胞的分布、表型、发育以及功能也有差别(表1-5)。
表1-5 TCRαβ与TCRγδ细胞特性的比较
| 特性 | TCRαβ | TCRγδ |
| |
| 分 布 | PBL | 60~70% | 0.5~15% |
|
| 其它部分 | 小鼠树突状表皮细胞(DEC)、小鼠粘膜上皮内淋巴细胞(IEL) |
| ||
| 表 型 | CD4+CD8- | 60~65% | <1% |
|
| CD4-CD8+ | 35% | 20~50% |
| |
| CD4-CD8- | <1% | 50~80% |
| |
| CD2 | 100% | 100% |
| |
| CD5 | >95% | -或+(弱) |
| |
| 发 育 | 胸腺后期 | 胸腺早期(早于TCRαβ) |
| |
| 功 能 | (1)识别MHC与加工处理 多肽的复合物 | (1)识别外来抗原的MHC限制尚有争论,识别破伤风类毒素可能受MHCⅡ类抗原限制;小鼠TCRγδ对合成肽反应受Qa分子限制。 |
| |
| (2)识别非已MHC及MHCⅠ类抗原相关分子(TLa、CD1) | ||||
| (2)识别非已MHC | ||||
| (3)产生些细胞因子(IL-2、IL-4、IL-5、GM-CSF、IFN-γ) |
| |||
| (3)产生多种淋巴因子 | (4)某些TCRγδ细胞可杀伤靶细胞 |
| ||
| (5)调节TCRαβ的发育。 | ||||
| (4)杀伤病毒感染等靶 细胞 | ||||
| (6)上皮屏障(针对肠毒素、分枝杆菌热休克蛋白等) |
| |||
注:树突状表皮细胞dendriticepidermal cell, DEC
上皮内淋巴细胞intraepithelial lymphocyte IEL
在正常外周血中,CD4-CD8+、CD4+CD8-、CD4+CD8+和CD4-CD8-四种表型不同的T细胞分别占T细胞总数的25%、70%、1%和4%左右,其中前三种表型TCR类型主要为TCRαβ,而CD4-CD8-T细胞主要为TCRγδ。以下疾病可见外周血或局部TCRγδ细胞数量或比率升高:(1)重症联合免疫缺陷、常见可变型免疫缺陷、Wiskott-Aldrich综合征、Di-George综合征、白血病患者骨髓移植等病人外周血中TCRγδ细胞百分率增加;(2)少数急性T细胞白血病、T细胞恶性淋巴瘤患者为TCRγδT细胞发生恶性变;(3)慢性淋巴细胞性白血病、大颗粒淋巴细胞(LGL)白血病病人PBMC中TCRγδT细胞百分率增加;(4)肾移植患者排异反应晚期外周血中TCRγδ细胞增加;(5)类风湿性关节炎患者关节腔滑液中TCRγδ阳性细胞比率要高于外周血中TCRγδ细胞比例,推测TCRγδ可能参与局部炎症的发生;(6)经结核杆菌免疫后的局部淋巴引流液中TCRγδT细胞比例增加,麻风结节病灶中有很高比例的TCRγδT细胞,提示TCRγδT细胞对分枝杆菌所致的感染免疫中起重要作用;(7)HIV、EBV感染时外周血TCRγδT细胞比例增加。
[超搞原]有的抗原不经过APC处理和递呈可直接激活CD4阳性T细胞称为超抗原(superantigen,SAg),具有类似致分裂原的作用。SAg对T细胞的激活采取一种独特的方式,即分子一端和TCRβ链上V基因产物结合,别一端和APC表面MHCⅡ类分子相结合。因此SAg发挥作用需有两类细胞:表达TCRβ链的CD4+T细胞和表达MHCⅡ类抗原的辅佐细胞。外源性的SAg主要是葡萄菌、链球菌、支原体等微生物产生的毒素,其中以葡萄球菌肠毒素A(Staphylococcus enterotoxin A,SEA)研究得最多,SEA往往取用特定TCR基因片段Vβ6.9及Vβ22,SEA另一侧与HLA-DR分子β1结构域的α螺旋相结合。
SAg激活CD4+T细胞使之释放IL-2,IFN、TNF等细胞因子,诱导CTL分化为效应细胞,可杀伤对NK、LAK高度抵抗的白血病细胞。
- T细胞受体《细胞和分子免疫学》
- T细胞缺陷性疾病《医学免疫学》
- T细胞受体基因《细胞和分子免疫学》
- T细胞旁路(t cell by - pass)学说《实用免疫细胞与核酸》
- T细胞亚群的分类及功能《医学免疫学》
- T细胞库的多样性《细胞和分子免疫学》
- T细胞在胸腺分化过程中的表型改变《细胞和分子免疫学》
- T细胞介导的细胞毒试验《免疫学和免疫学检验》
- T细胞在胸腺内的发育《医学免疫学》
- T细胞基因的转录激活及其表达《细胞和分子免疫学》
- T细胞在胸腺中的选择《细胞和分子免疫学》
- T细胞功能的检测《免疫学和免疫学检验》
- T细胞在胸腺中获得MHC限制的能力《细胞和分子免疫学》
- T细胞表面标志的检测《免疫学和免疫学检验》
- T细胞增殖试验《免疫学和免疫学检验》
- T细胞《医学免疫学》
- T细胞主要表面分子《医学免疫学》
- T细胞《细胞和分子免疫学》
- t值表《预防医学》
- T细胞《免疫学和免疫学检验》
- U《医学统计学》
- T淋巴细胞亚群《细胞和分子免疫学》
- U《医学遗传学基础》
- T淋巴细胞及其亚群的免疫表达《实用免疫细胞与核酸》
- U《传染病》
- T淋巴细胞及其亚群《细胞和分子免疫学》
- USG的成像基本原理与设备《医学影像学》
- T淋巴细胞分布及其特点《实用免疫细胞与核酸》
- USG检查技术《医学影像学》
- T淋巴细胞《实用免疫细胞与核酸》
- USG论断《医学影像学》